
 |
| Table: Classification of known CoV |
Most CoV identified
until now are causing severe disease only in animals including agricultural
important animals such as chicken, cattle, and pigs. To date only six human CoV
(HCoV) have been identified, namely HCoV-229E, HCoV-OC43, HCoV-NL63, HCoV-HKU1,
Severe Respiratory Syndrome (SARS)-CoV and most recently Middle Eastern
Respiratory Syndrome (MERS)-CoV, although a SARS-like CoV, WIV1-CoV, replicates
in primary human epithelial cells at low levels as well as in mice expressing
the human SARS-CoV receptor ACE2 albeit at lower levels compared to
SARS-CoV.
In contrast to
SARS-CoV and MERS-CoV, the infection with HCoV-229E, HCoV-OC43, HCoV-NL63 and
HCoV-HKU1only causes mild upper respiratory tract infections, whereas the
infection with MERS-CoV causes severe lower respiratory infection and renal
failure which may lead to the death of infected individuals.
Although the natural
host for MERS-CoV has not been conclusively identified, bats might act as a
natural reservoir of MERS-CoV (similar to Hendra and Nipah Virus) as evidenced
by studies indicating the presence of MERS-like CoV in bats from Saudi Arabia,
Europe, Africa and Asia. Furthermore, MERS-CoV can enter and replicate both in
cell lines derived from bats and humans and the closely related Bat CoV-HKU4
can enter cells by binding to DPP4, the receptor for MERS-CoV. In Jamaican
fruit bats (Artibeus jamaicensis)
experimentally infected with MERS-CoV, clinical signs are absent despite viral
replication and shedding in the respiratory and intestinal tract for up to 9
days p.i. as well as the induction of an antiviral response (see below for
discussion).
Despite the ability of
MERS and MERS-like CoV to infect both human and bat cell lines, bats are
unlikely to be the source for the current outbreak of MERS in the Arabian
Peninsula based on findings that livestock –in particular dromedary camels-
have a high seroprevalence for MERS-CoV. In addition, recent studies showed
that DPP4 is expressed at high levels in the upper respiratory tract of
dromedary camels (but not humans) and that sequences of MERS-CoV isolated from
dromedary camels are highly similar to sequence from human derived viral
isolates. Finally, camel derived MERS-CoV can efficiently replicate in human
cells.
MERS-CoV
and the Interferon response
The polycistronic
single stranded positive sense viral RNA of MERS-CoV encodes for two
polyproteins, orf1a and orf1b, within the 5’ end which are cleaved into a total
of 16 non-structural proteins (nsp) including the viral RNA dependent RNA
polymerase whereas the 3’ end of the viral RNA encodes for the structural
proteins, namely spike (S), membrane (M), envelope (E) protein as well as
lineage specific proteins.
 |
| Figure: Processing of the orf1ab into several nsp |
Following viral entry
and release of the viral RNA, MERS-CoV replicates in the cytoplasm of cell,
although the viral N protein is imported into the nucleus and localizes to the
nucleolus. During viral replication, replication centers containing viral
proteins and dsRNA are formed in the ER-Golgi intermediate compartment (ERGIC)
in a process that might involve the induction of autophagy and/or viral induced
rearrangements of the ER membrane that promote the formation of vesicular
structures.
 |
| Figure: Outline of CoV replication cycle |
The infection of a
variety of cell lines with SARS-CoV and MERS-CoV has been reported to induce
the expression of type I interferons (IFN) including IFN-α and IFN-β,
suggesting that both SARS-CoV and MERS-CoV induce antiviral signaling via the
recognition of viral RNA by PRR and PAMPS. Furthermore, both SARS-CoV and
MERS-CoV express proteins that counteract the IFN pathway and thus contribute
to the severity of infection. In general, MERS-CoV RNA is recognized by viral
sensors of the retinoic acid-inducible gene-I (RIG-1)-like receptor (RLR)
family melanoma differentiation gene 5 (MDA5) that upon activation by viral
dsRNA intermediates, induce the nuclear translocation of both IFN regulatory
factor (IRF)-3 and nuclear factor κB (NF-κB), both of which induce the
expression of IFN-β. RIG-1 signaling
induced by the viral RNA as well as by Poly (I:C) however can be inhibited by
several viral proteins, including ORF4a, ORF4b, ORF5 and M. In the case of
ORF4a, the viral protein binds dsRNA and thus inhibits the activation of RIG-1
and MDA-5, whereas the nuclear localization of ORF4b inhibits the induction of
IFN-β as well as the expression of RIG-I, MDA5, MAVS, IKKε, and TBK-1 by
inhibiting both (nuclear) IRF-3 and IRF-7 whereas in the cytoplasm ORF4b
inhibits antiviral signaling by binding to TANK binding kinase 1 (TBK1) and IκB
kinase epsilon (IKK-ε) thus suppressing the interaction between mitochondrial
antiviral signaling protein (MAVS) and IKK-ε. In addition to inhibiting IRF-3
by binding to TBK1, both MERS-CoV and ORF4b protein also inhibits antiviral
signaling by inhibiting the IFN inducible oligoadenylate synthetase (OAS)-RNase
L pathway that is activated by the viral dsRNA intermediate via degradation of 2′,5′-oligoadenylate
(2-5A), the activator of RNase L, thus inhibiting RIG-1 and MDA-5 induced
signaling. Interestingly, ORF4b localises to the nucleolus in ORF4b transfected
cells, but so far the importance for nucleolar localisation of ORF4b for
antiviral signaling has not been demonstrated. In contrast to ORF4b, ORF5 –like
ORF4a and M- can only be detected in the cytoplasm of infected cells. Similar
to ORF4b however, the expression of ORF5 inhibits the induction of IFN-β upon
infection with Sendai Virus (albeit to a lesser effect) probably by inhibiting
IRF-3 and -7 but not NF-κB.
Activated
RNaseL cleaves viral and cellular ssRNA including ribosomal RNA preferentially
at UU and UA dinucleotide residues, therefore not only inhibiting replication
of the viral genome but also the expression of viral proteins as well as
processing viral RNA to be recognized by intracellular RNA sensing proteins
thus activating antiviral signaling pathways.
In the case of CoV, the Mouse Hepatitis Virus
(MHV) NS2 protein encoded by ORF2a has been identified as an inhibitor of the OAS-RNaseL
pathway by inhibiting the conversion of intracellular ATP to 2’,5’- OAS via the
2’,5’ Phosphodiesterase domain (PDE) following the detection of viral dsRNA
intermediates or synthetic Poly (I:C) RNA. Alignment of MERS-CoV, BtCoV-HKU5
and BtCoV-SC2013 orf4b with MHV nsp2 lead to the identification of a 2’,5’- PDE
domain that is preceded by a nuclear localization signal that is catalytically
active in a fluorescence resonance energy transfer (FRET) based RNaseL activity
assay with similar kinetics to MHV nsp2. As expected, all orf4b proteins
localize to the nucleus and the cytoplasm if expressed in murine L2 cells
infected with MHV chimeras that express a catalytically inactive mutant of nsp2
and catalytically active orf4b derived from MERS-CoV or BatCoV-SC2013; in case
of the MHV-BtCoV-HKU5 orf4b chimera, orf4b predominantly localises to the
nucleus which does not affect viral
replication but might not inhibit the activation of RNaseL, indicating that
only cytoplasmic orf4b can inhibit RNaseL activation. In Calu-3 cells, a human
airway epithelial cell line, infected with MERS-CoV, the deletion of nsp3-5 or
the deletion of orf4b does not activate the degradation of rRNA to the full
extent when compared to wt MERS-CoV probably due to the alternative pathways
that inhibit OAS dependent activation of RNAseL such as the orf4a dependent
sequestering of viral dsRNA intermediates.
In addition to the localisation of orf4b, the presence of a second His residue within the PDE is important for the ability of orf4b to regulate the activity of RNaseL since substitution of the H182 (MERS-CoV) or H 186 (BtCoV SC2013) with R result in decreased ability of orf4b to inhibit RNaseL.
In addition to the localisation of orf4b, the presence of a second His residue within the PDE is important for the ability of orf4b to regulate the activity of RNaseL since substitution of the H182 (MERS-CoV) or H 186 (BtCoV SC2013) with R result in decreased ability of orf4b to inhibit RNaseL.
 |
| Figure: N-terminal NLS in orf4b is required for nuclear localisation of MERS-Cov and BtCoV off 4b |
 |
| Figure: Cytoplasmic localisation and His at position of 182 or 186 of MERS-CoV and BtCoV orf4b are required for inhibition of RNaseL |
The induction of
antiviral signaling by via OAS/RNase L dependent pathways has also been
demonstrated for West Nile Virus (WNV), Influenza A Virus (IAV) and Vaccinia
Virus (VACV). OAS3 knockout cells infected with WNV, IAV, VACV or treated with
Poly(I:C) exhibit only minimal 2’-5’ oligoadenylate synthase activity and
infected A549, hTERT-HME and HT1080 OAS3 knockout cells display higher viral
titres than the corresponding wt cells. Interestingly, probably because of
increased expression of IFN-λ1, trophoblasts of the human placenta express
higher levels of OAS-1,-2, and -3 compared to JEG-3 cells, suggesting that
increased OAS levels contribute to the resistance of trophoblasts to ZIKV
resistance as well.
 |
| Figure: orf4b degrades 2',5' OAS and prevents activation of RNaseL |
More recently, the
viral M protein of both SARS-CoV and MERS-CoV but not HCoV-HKU1 has been
demonstrated to form a complex with the TRAF3 adapter protein, thus inhibiting
the formation of the TRAF3-TANK-TBK1/ IKK-ε complex via the N terminal domain
of the viral M protein. Consequently, the expression of MERS-CoV or SARS-CoV
inhibits RLR signaling induced by Sendai Virus or Poly (I:C) and further
analysis using RIG-1 and NF-κB reporter plasmids showed that the expression of
MERS-COV M preferentially inhibits RIG-1 mediated activation of IRF-3, thus
preventing the induction of IFN-β. Similar to SARS-CoV M, the interaction
between TRAF3 and MERS-CoV M is mediated by the N-terminal transmembrane domain
1 whereas the two other transmembrane domains as well as the C-terminal domain
are dispensable. Both the SARS-CoV and MERS-CoV derived M and ORF 4b protein
therefore inhibit antiviral RIG-1 by targeting the formation of the
TRAF3-TANK-TBK1/ IKK-ε complex, thus inhibiting IRF-3 dependent signaling. From
an experimental standpoint it would be interesting if a recombinant HCoV-HKU1
expressing MERS-CoV or SARS-CoV derived M exhibits increased viral titres
following infection of primary human cells compared to wt virus. Also, it
remains to be seen if the expression of MERS-CoV in Artibeus jamaicensis or in bat derived cell lines inhibits the
antiviral signaling in a similar way. So far, the infection of Artibeus jamaicensis with MERS-CoV has
been demonstrated to increase the expression of Mx-1, ISG56 and RANTES
moderately suggesting that MERS-CoV does inhibit antiviral signaling in bats,
thus explaining the absence of clinical signs in infected bats despite the
presence of low viral titres in the duodenum of bats. The individual role of
ORF4a, ORF4b, ORF5 and M has not been assessed in bat or camel derived cell
lines. Indeed, the importance of the genetic and functional diversity of
proteins involved in the induction of antiviral signaling pathways has recently
been highlighted in studies comparing RIG-1 from waterfowl and mammals in their
capacity to facilitate an immune response to Influenza A virus. In short, duck
derived RIG-1 had a weaker ability to induce IFN compared to goose derived
RIG-1 and both duck and goose derived RIG-1 are more effective than pigeon
RIG-1 in (chicken) DF-1 cells infected with Influenza A virus.
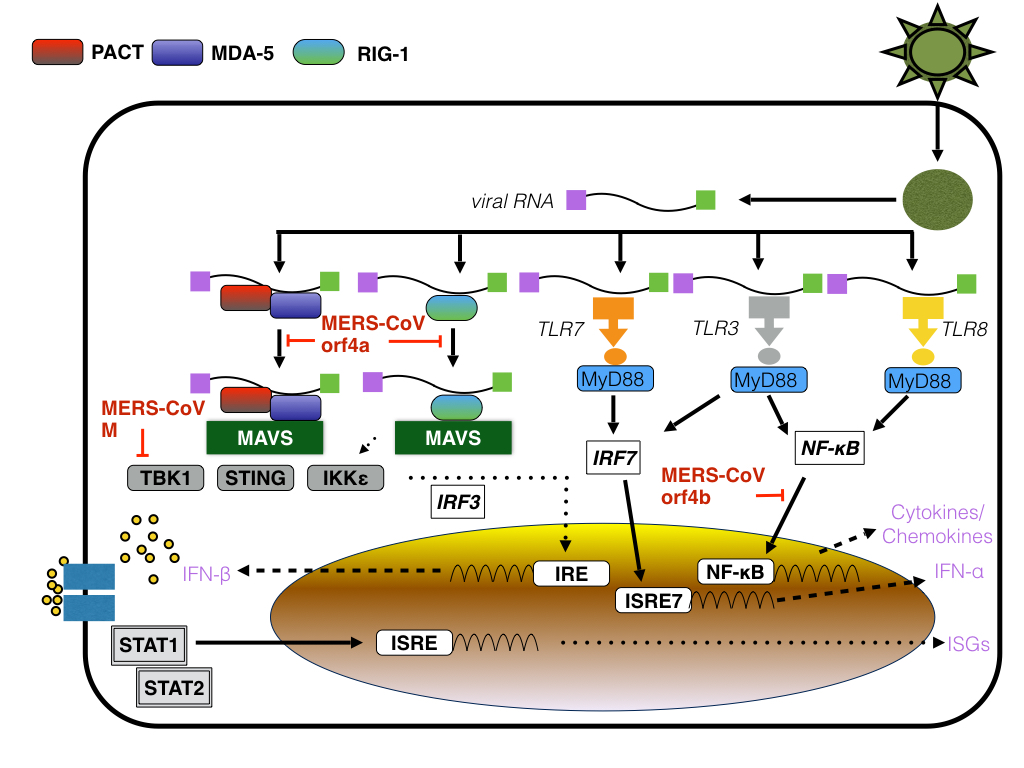 |
| Figure: MERS-CoV M, orf4a and orf4b: points of action |
So far it has not been
shown if the expression of mutant orf4b in the context of Coronavirus infection
not only abrogates immune signaling and thus promotes viral replication solely
by inhibiting apoptosis but also abrogates the induction of RNaseL induced
autophagy and degradation of viral replication centers. If so, the number of
viral RC should be increased in cells expressing mutant orf4b and/or mouse
embryonic fibroblasts derived from RNaseL -/- mice.
Further reading
Han HJ, Yu H, & Yu XJ (2016). Evidence for zoonotic origins of Middle East respiratory syndrome coronavirus. The Journal of general virology, 97 (2), 274-80 PMID: 26572912
Han, H., Wen, H., Zhou, C., Chen, F., Luo, L., Liu, J., & Yu, X. (2015). Bats as reservoirs of severe emerging infectious diseases Virus Research, 205, 1-6 DOI: 10.1016/j.virusres.2015.05.006
Chan JF, Lau SK, To KK, Cheng VC, Woo PC, & Yuen KY (2015). Middle East respiratory syndrome coronavirus: another zoonotic betacoronavirus causing SARS-like disease. Clinical microbiology reviews, 28 (2), 465-522 PMID: 25810418
Wong LY, Lui PY, & Jin DY (2016). A molecular arms race between host innate antiviral response and emerging human coronaviruses. Virologica Sinica, 31 (1), 12-23 PMID: 26786772
Lin R, Heylbroeck C, Pitha PM, & Hiscott J (1998). Virus-dependent phosphorylation of the IRF-3 transcription factor regulates nuclear translocation, transactivation potential, and proteasome-mediated degradation. Molecular and cellular biology, 18 (5), 2986-96 PMID: 9566918
Yang XL, Hu B, Wang B, Wang MN, Zhang Q, Zhang W, Wu LJ, Ge XY, Zhang YZ, Daszak P, Wang LF, & Shi ZL (2015). Isolation and Characterization of a Novel Bat Coronavirus Closely Related to the Direct Progenitor of Severe Acute Respiratory Syndrome Coronavirus. Journal of virology, 90 (6), 3253-6 PMID: 26719272
Menachery VD, Yount BL Jr, Sims AC, Debbink K, Agnihothram SS, Gralinski LE, Graham RL, Scobey T, Plante JA, Royal SR, Swanstrom J, Sheahan TP, Pickles RJ, Corti D, Randell SH, Lanzavecchia A, Marasco WA, & Baric RS (2016). SARS-like WIV1-CoV poised for human emergence. Proceedings of the National Academy of Sciences of the United States of America PMID: 26976607
Munster, V., Adney, D., van Doremalen, N., Brown, V., Miazgowicz, K., Milne-Price, S., Bushmaker, T., Rosenke, R., Scott, D., Hawkinson, A., de Wit, E., Schountz, T., & Bowen, R. (2016). Replication and shedding of MERS-CoV in Jamaican fruit bats (Artibeus jamaicensis) Scientific Reports, 6 DOI: 10.1038/srep21878
Yang Y, Ye F, Zhu N, Wang W, Deng Y, Zhao Z, & Tan W (2015). Middle East respiratory syndrome coronavirus ORF4b protein inhibits type I interferon production through both cytoplasmic and nuclear targets. Scientific reports, 5 PMID: 26631542
Siu KL, Kok KH, Ng MH, Poon VK, Yuen KY, Zheng BJ, & Jin DY (2009). Severe acute respiratory syndrome coronavirus M protein inhibits type I interferon production by impeding the formation of TRAF3.TANK.TBK1/IKKepsilon complex. The Journal of biological chemistry, 284 (24), 16202-9 PMID: 19380580
Matthews, K., Schäfer, A., Pham, A., & Frieman, M. (2014). The SARS coronavirus papain like protease can inhibit IRF3 at a post activation step that requires deubiquitination activity Virology Journal, 11 (1) DOI: 10.1186/s12985-014-0209-9
Siu, K., Yeung, M., Kok, K., Yuen, K., Kew, C., Lui, P., Chan, C., Tse, H., Woo, P., Yuen, K., & Jin, D. (2014). Middle East Respiratory Syndrome Coronavirus 4a Protein Is a Double-Stranded RNA-Binding Protein That Suppresses PACT-Induced Activation of RIG-I and MDA5 in the Innate Antiviral Response Journal of Virology, 88 (9), 4866-4876 DOI: 10.1128/JVI.03649-13
Lui PY, Wong LY, Fung CL, Siu KL, Yeung ML, Yuen KS, Chan CP, Woo PC, Yuen KY, & Jin DY (2016). Middle East respiratory syndrome coronavirus M protein suppresses type I interferon expression through the inhibition of TBK1-dependent phosphorylation of IRF3. Emerging microbes & infections, 5 PMID: 27094905
Matthews KL, Coleman CM, van der Meer Y, Snijder EJ, & Frieman MB (2014). The ORF4b-encoded accessory proteins of Middle East respiratory syndrome coronavirus and two related bat coronaviruses localize to the nucleus and inhibit innate immune signalling. The Journal of general virology, 95 (Pt 4), 874-82 PMID: 24443473
Thornbrough JM, Jha BK, Yount B, Goldstein SA, Li Y, Elliott R, Sims AC, Baric RS, Silverman RH, & Weiss SR (2016). Middle East Respiratory Syndrome Coronavirus NS4b Protein Inhibits Host RNase L Activation. mBio, 7 (2) PMID: 27025250
Li Y, Banerjee S, Wang Y, Goldstein SA, Dong B, Gaughan C, Silverman RH, & Weiss SR (2016). Activation of RNase L is dependent on OAS3 expression during infection with diverse human viruses. Proceedings of the National Academy of Sciences of the United States of America, 113 (8), 2241-6 PMID: 26858407
Banerjee S, Chakrabarti A, Jha BK, Weiss SR, & Silverman RH (2014). Cell-type-specific effects of RNase L on viral induction of beta interferon. mBio, 5 (2) PMID: 24570368
Castelli, J., Wood, K., & Youle, R. (1998). The 2-5A system in viral infection and apoptosis Biomedicine & Pharmacotherapy, 52 (9), 386-390 DOI: 10.1016/S0753-3322(99)80006-7
Malathi, K., Dong, B., Gale, M., & Silverman, R. (2007). Small self-RNA generated by RNase L amplifies antiviral innate immunity Nature, 448 (7155), 816-819 DOI: 10.1038/nature06042
Shao Q, Xu W, Guo Q, Yan L, Rui L, Liu J, Zhao Y, & Li Z (2015). RIG-I from waterfowl and mammals differ in their abilities to induce antiviral responses against influenza A viruses. The Journal of general virology, 96 (Pt 2), 277-87 PMID: 25371516
Xu W, Shao Q, Zang Y, Guo Q, Zhang Y, & Li Z (2015). Pigeon RIG-I Function in Innate Immunity against H9N2 IAV and IBDV. Viruses, 7 (7), 4131-51 PMID: 26205406